Взаимодействие аллельных и неаллельных генов. Явление плейотропии
Генотип — не просто механический набор генов, это исторически сложившаяся система из взаимодействующих между собой генов. Точнее, взаимодействуют не сами гены (участки молекул ДНК), а образуемые на их основе продукты (РНК и белки).
Взаимодействовать могут как аллельные гены, так и неаллельные.
| Тип взаимодействия генов | Характер взаимодействия | Расщепление по фенотипу в F 2 | Генотипический состав фенотипических классов | Пример | |
|---|---|---|---|---|---|
| Взаимодействие аллельных генов | |||||
| Полное доминирование | Доминантный аллель A подавляет рецессивный аллель a | 3:1 | 3A- : 1aa | Наследование цвета семян гороха | |
| Неполное доминирование | Признак у гетерозиготной формы выражен слабее, чем у гомозиготной | 1:2:1 | 1AA: 2Aa: 1aa | Наследование окраски цветков ночной красавицы | |
| Кодоминирование | В гетерозиготном состоянии каждый из аллельных генов вызывает развитие контролируемого им признака | 1:2:1 | 1I A I A: 2I A I B: 1I B I B | Наследование групп крови у человека | |
| Взаимодействие неаллельных генов | |||||
| Кооперация | Доминантные гены из разных пар (A, B), присутствуя в генотипе вместе, вызывают формирование нового признака. Присутствуя каждый по отдельности, гены А и В вызывают развитие своих признаков | 9:3:3:1 | 9A-B- : 3A-bb: 3aaB- : 1aabb | Наследование формы гребня кур | |
| Комплементарность | Доминантные гены из разных пар (A, B), присутствуя в генотипе вместе, вызывают формирование нового признака. Присутствуя каждый по отдельности, гены A и B развитие признака не вызывают | 9:7 | (9A-B-) : (3A-BB + 3aaB- + 1aabb) | Наследование цвета цветков душистого горошка | |
| Эпистаз | доминантный | Гены одной аллельной пары подавляют действие генов другой | 13:3 | (9I-C- + 3I-cc + 1iicc) : (3cci-) | Наследование окраски оперения кур |
| рецессивный | 9:3:4 | 9A-C- : 3aaC- : (3A-cc + 1aacc) | Наследование окраски шерсти у домовых мышей | ||
| Полимерия | Одновременное действие нескольких неаллельных генов | 15:1 | (9A 1 -A 2 + 3A 1 -a 2 a 2 + 3a 1 a 1 A 2 -) : 1a 1 a 1 a 2 a 2 | Наследование цвета кожи у человека | |
Взаимодействие аллельных генов
Различают три типа взаимодействия аллельных генов: полное доминирование, неполное доминирование, кодоминирование.
- Полное доминирование — явление, когда доминантный ген полностью подавляет работу рецессивного гена, в результате чего развивается доминантный признак.
- Неполное доминирование — явление, когда доминантный ген не полностью подавляет работу рецессивного гена, в результате чего развивается промежуточный признак.
- Кодоминирование (независимое проявление) — явление, когда в формировании признака у гетерозиготного организма участвуют оба аллеля. У человека с серией множественных аллелей представлен ген, определяющий группу крови. При этом гены, обусловливающие группы крови A и B, являются кодоминантными по отношению друг к другу и оба доминантны по отношению к гену, определяющему группу крови 0.
Взаимодействие неаллельных генов
Различают четыре типа взаимодействия неалльлельных генов: кооперацию, комплементарность, эпистаз и полимерию.
Кооперация — явление, когда при взаимном действии двух доминантных неаллельных генов, каждый из которых имеет свое собственное фенотипическое проявление, происходит формирование нового признака.
Комплементарность — явление, когда признак развивается только при взаимном действии двух доминантных неаллельных генов, каждый из которых в отдельности не вызывает развитие признака.
Эпистаз — явление, когда один ген (как доминантный, так и рецессивный) подавляет действие другого (неаллельного) гена (как доминантного, так и рецессивного). Ген-подавитель (супрессор) может быть доминантным (доминантный эпистаз) или рецессивным (рецессивный эпистаз).
Полимерия — явление, когда несколько неаллельных доминантных генов отвечают за сходное воздействие на развитие одного и того же признака. Чем больше таких генов присутствуют в генотипе, тем ярче проявляется признак. Явление полимерии наблюдается при наследовании количественных признаков (цвет кожи, масса тела, удой коров).
В противоположность полимерии наблюдается такое явление, как плейотропия — множественное действие гена, когда один ген отвечает за развитие нескольких признаков.
Теперь обратимся к проблеме взаимодействия неаллельных генов. Если развитие признака контролируется более чем одной парой генов, то это означает, что он находится под полигенным контролем. Установлено несколько основных типов взаимодействия генов: комплементарность, эпистаз, полимерия и плейотропия.
Первый случай неаллельного взаимодействия был описан в качестве примера отклонения от законов Менделя английскими учеными У. Бетсоном и Р. Пеннетом в 1904 г. при изучении наследования формы гребня у кур. Различные породы кур характеризуются разной формой гребня. Виандотты имеют низкий, правильный, покрытый сосочками гребень, известный под названием “розовидного”. Брамы и некоторые бойцовые куры обладают узким и высоким гребнем с тремя продольными возвышениями — “гороховидным”. Леггорны имеют простой или листовидный гребень, состоящий из одной вертикальной пластинки. Гибридологический анализ показал, что простой гребень ведет себя как полностью рецессивный признак по отношению к розовидному и гороховидному. Расщепление в F 2 соответствует формуле 3: 1. При скрещивании же между собой рас с розовидным и гороховидным гребнем у гибридов первого поколения развивается совершенно новая форма гребня, напоминающая половинку ядра грецкого ореха, в связи с чем гребень был назван “ореховидным”. При анализе второго поколения было установлено, что соотношение разных форм гребня в F 2 соответствует формуле 9: 3: 3: 1, что указывало на дигибридный характер скрещивания. Была разработана схема скрещивания, объясняющая механизм наследования этого признака.
В определении формы гребня у кур принимают участие два неаллельных гена. Доминантный ген R контролирует развитие розовидного гребня, а доминантный ген P — гороховидного. Комбинация рецессивных аллелей этих генов rrpp вызывает развитие простого гребня. Ореховидный гребень развивается при наличии в генотипе обоих доминантных генов.
Наследование формы гребня у кур можно отнести к комплементарному взаимодействию неаллельных генов. Комплементарными, или дополнительными, считаются гены, которые при совместном действии в генотипе в гомо- или гетерозиготном состоянии обусловливают развитие нового признака. Действие же каждого из генов в отдельности воспроизводит признак одного из родителей.
Схема, иллюстрирующая взаимодействие неаллельных генов,
определяющих форму гребня у кур
Наследование генов, определяющих форму гребня у кур, полностью укладывается в схему дигибридного скрещивания, так как они ведут себя при распределении независимо. Отличие от обычного дигибридного скрещивания проявляется только на уровне фенотипа и сводится к следующему:
- Гибриды F 1 не похожи ни на одного из родителей и обладают новым признаком;
- В F 2 появляются два новых фенотипических класса, которые являются результатом взаимодействия либо доминантных (ореховидный гребень), либо рецессивных (простой гребень) аллелей двух независимых генов.
Механизм комплементарного взаимодействия подробно изучен на примере наследования окраски глаз у дрозофилы. Красная окраска глаз у мух дикого типа определяется одновременным синтезом двух пигментов — бурого и ярко-красного, каждый из которых контролируется доминантным геном. Мутации, затрагивающие структуру этих генов, блокируют синтез либо того, либо другого пигмента. Так, рецессивная мутация brown (ген находится во 2-й хромосоме) блокирует синтез ярко-красного пигмента, в связи с чем у гомозигот по этой мутации бурые глаза. Рецессивная мутация scarlet (ген располагается в 3-й хромосоме) нарушает синтез бурого пигмента, и поэтому гомозиготы stst имеют ярко-красные глаза. При одновременном присутствии в генотипе обоих мутантных генов в гомозиготном состоянии не вырабатываются оба пигмента и глаза у мух белые.
В описанных примерах комплементарного взаимодействия неаллельных генов формула расщепления по фенотипу в F 2 соответствует 9: 3: 3: 1. Такое расщепление наблюдается в том случае, если взаимодействующие гены по отдельности имеют неодинаковое фенотипическое проявление и оно не совпадает с фенотипом гомозиготного рецессива. Если это условие не соблюдается, в F 2 имеют место иные соотношения фенотипов.
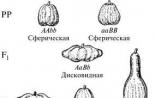
Например, при скрещивании двух разновидностей фигурной тыквы со сферической формой плода гибриды первого поколения обладают новым признаком — плоскими или дисковидными плодами. При скрещивании гибридов между собой в F 2 наблюдается расщепление в соотношении 9 дисковидных: 6 сферических: 1 удлиненная.

Анализ схемы показывает, что в определении формы плода принимают участие два неаллельных гена с одинаковым фенотипическим проявлением (сферическая форма). Взаимодействие доминантных аллелей этих генов дает дисковидную форму, взаимодействие рецессивных аллелей — удлиненную.
Еще один пример комплементарного взаимодействия дает наследование окраски шерсти у мышей. Дикая серая окраска определяется взаимодействием двух доминантных генов. Ген А отвечает за присутствие пигмента, а ген В — за его неравномерное распределение. Если в генотипе присутствует только ген А (А-bb ), то мыши равномерно окрашены в черный цвет. Если присутствует только ген В (ааВ- ), то пигмент не вырабатывается и мыши оказываются неокрашенными, так же как и гомозиготный рецессив ааbb . Такое действие генов приводит к тому, что в F 2 расщепление по фенотипу соответствует формуле 9: 3: 4.

F 2
| AB | Ab | aB | ab | |
| AB | AABB сер. |
AABb сер. |
AaBB сер. |
AaBb сер. |
| Ab | AABb сер. |
AAbb черн. |
AaBb сер. |
Aabb черн. |
| aB | AaBB сер. |
AaBb сер. |
aaBB бел. |
aaBb бел. |
| ab | AaBb сер. |
Aabb черн. |
aaBb бел. |
aabb |
Комплементарное взаимодействие описано также при наследовании окраски цветов у душистого горошка. Большая часть сортов этого растения имеет пурпурные цветы с фиолетовыми крыльями, которые характерны для дикой сицилийской расы, но есть также сорта с белой окраской. Скрещивая растения с пурпурной окраской цветов с растениями с белыми цветами Бетсон и Пеннет установили, что пурпурная окраска цветов полностью доминирует над белой, и в F 2 наблюдается соотношение 3: 1. Но в одном случае от скрещивания двух белых растений получилось потомство, состоящее только из растений с окрашенными цветами. При самоопылении растений F 1 было получено потомство, состоящее из двух фенотипических классов: с окрашенными и неокрашенными цветами в соотношении 9/16: 7/16.
Полученные результаты объясняются комплементарным взаимодействием двух пар неаллельных генов, доминантные аллели которых (С и Р ) в отдельности не способны обеспечить развитие пурпурной окраски, так же как и их рецессивные аллели (ссрр ). Окраска проявляется только при наличии в генотипе обоих доминантных генов, взаимодействие которых обеспечивает синтез пигмента.

пурп.
F 2
| CP | Cp | cP | cp | |
| CP | CCPP пурп. |
CCPp пурп. |
CcPP пурп. |
CcPp пурп. |
| Cp | CCPp пурп. |
CCpp бел. |
CcPp пурп. |
Ccpp бел. |
| cP | CcPP пурп. |
CcPp пурп. |
ccPP бел. |
ccPp бел. |
| cp | CcPp пурп. |
Ccpp бел. |
ccPp бел. |
В приведенном примере формула расщепления в F 2 — 9: 7 обусловлена отсутствием у доминантных аллелей обоих генов собственного фенотипического проявления. Однако такой же результат получается и в том случае, если взаимодействующие доминантные гены имеют одинаковое фенотипическое проявление. Например, при скрещивании двух сортов кукурузы с фиолетовой окраской зерновок в F 1 все гибриды имеют желтые зерновки, а в F 2 наблюдается расщепление 9/16 желт. : 7/16 фиол.
Эпистаз — другой тип неаллельного взаимодействия, при котором происходит подавление действия одного гена другим неаллельным ему геном. Ген, который препятствует проявлению другого гена, называется эпистатичным, или супрессором, а тот, чье действие подавляется, гипостатичным. В качестве эпистатичного гена может выступать как доминантный, так и рецессивный ген (соответственно доминантный и рецессивный эпистаз).
Примером доминантного эпистаза служит наследование окраски шерсти у лошадей и окраски плодов у тыквы. Схема наследования этих двух признаков абсолютно одинаковая.

F 2
| CB | Cb | cB | cb | |
| CB | CCBB сер. |
CCBB сер. |
CcBB сер. |
CcBb сер. |
| Cb | CCBb сер. |
CCbb сер. |
CcBb сер. |
Ccbb сер. |
| cB | CcBB сер. |
CcBb сер. |
ccBB черн. |
ccBb черн. |
| cb | CcBb сер. |
Ccbb сер. |
ccBb черн. |
ccbb рыж. |
Из схемы видно, что доминантный ген серой окраски С является эпистатичным по отношению к доминантному гену В , который обусловливает черную окраску. В присутствии гена С ген В своего действия не проявляет, и поэтому гибриды F 1 несут признак, определяемый эпистатичным геном. В F 2 класс с обоими доминантными генами сливается по фенотипу (серая окраска) с классом, у которого представлен только эпистатичный ген (12/16). Черная окраска проявляется у 3/16 гибридных потомков, в генотипе которых отсутствует эпистатичный ген. В случае гомозиготного рецессива отсутствие гена-супрессора позволяет проявиться рецессивному гену с, который вызывает развитие рыжей окраски.

Доминантный эпистаз описан также при наследовании окраски пера у кур. Белый цвет оперенья у кур породы леггорнов доминирует над окрашенным черных, рябых и других цветных пород. Однако белая окраска других пород (например, плимутроков) рецессивна по отношению к цветному оперению. Скрещивания между особями с доминантной белой окраской и особями с рецессивной белой окраской в F 1 дают белое потомство. В F 2 наблюдается расщепление в соотношении 13: 3.
Анализ схемы показывает, что в определении окраски пера у кур принимают участие две пары неаллельных генов. Доминантный ген одной пары (I ) является эпистатичным по отношению к доминантному гену другой пары, вызывающему развитие окраски (C ). В связи с этим окрашенное оперение имеют только те особи, в генотипе которых присутствует ген С , но отсутствует эпистатичный ген I . У рецессивных гомозигот ссii отсутствует эпистатичный ген, но у них нет гена, который обеспечивает выработку пигмента (C ), поэтому они имеют белую окраску.
В качестве примера рецессивного эпистаза можно рассмотреть ситуацию с геном альбинизма у животных (см. выше схему наследования окраски шерсти у мышей). Присутствие в генотипе двух аллелей гена альбинизма (аа ) не дает возможности проявиться доминантному гену окраски (B ) — генотипы ааВ- .
Полимерный тип взаимодействия был впервые установлен Г. Нильсеном-Эле при изучении наследования окраски зерна у пшеницы. При скрещивании краснозерного сорта пшеницы с белозерным в первом поколении гибриды были окрашенными, но окраска была розовой. Во втором поколении только 1/16 часть потомства имела красную окраску зерна и 1/16 — белую, у остальных окраска была промежуточной с разной степенью выраженности признака (от бледно-розовой до темно-розовой). Анализ расщепления в F 2 показал, что в определении окраски зерна участвуют две пары неаллельных генов, действие которых суммируется. Степень выраженности красной окраски зависит от количества доминантных генов в генотипе.
Полимерные гены принято обозначать одинаковыми буквами с добавлением индексов, в соответствии с числом неаллельных генов.
Действие доминантных генов в данном скрещивании является аддитивным, так как добавление любого из них усиливает развитие признака.

F 2
| A 1 A 2 | A 1 a 2 | a 1 A 2 | a 1 a 2 | |
| A 1 A 2 | A 1 A 1 A 2 A 2 красн. |
A 1 A 1 A 2 Aa 2 ярко-розов. |
A 1 a 1 A 2 A 2 ярко-розов. |
A 1 a 1 A 2 a 2 розов. |
| A 1 a 2 | A 1 A 1 A 2 a 2 ярко-розов. |
A 1 A 1 a 2 a 2 розов. |
A 1 a 1 A 2 a 2 розов. |
A 1 a 1 a 2 a 2 бледно-розов. |
| a 1 A 2 | A 1 a 1 A 2 A 2 ярко-розов. |
A 1 a 1 A 2 a 2 розов. |
a 1 a 1 A 2 A 2 розов. |
a 1 a 1 A 2 a 2 бледно-розов. |
| a 1 a 2 | A 1 a 1 A 2 a 2 розов. |
A 1 a 1 a 2 a 2 бледно-розов. |
a 1 a 1 A 2 a 2 бледно-розов. |
a 1 a 1 a 2 a 2 |
Описанный тип полимерии, при котором степень развития признака зависит от дозы доминантного гена, называется кумулятивным. Такой характер наследования обычен для количественных признаков, к которым следует отнести и окраску, т.к. ее интенсивность обусловлена количеством вырабатываемого пигмента. Если не учитывать степень выраженности окраски, то соотношение окрашенных и неокрашенных растений в F 2 соответствует формуле 15: 1.
Однако в некоторых случаях полимерия не сопровождается кумулятивным эффектом. В качестве примера можно привести наследование формы семян у пастушьей сумки. Скрещивание двух рас, одна из которых имеет треугольные плоды, а другая яйцевидные дает в первом поколении гибриды с треугольной формой плода, а во втором поколении наблюдается расщепление по этим двум признакам в соотношении 15 треуг. : 1 яйцев.
Данный случай наследования отличается от предыдущего только на фенотипическом уровне: отсутствие кумулятивного эффекта при увеличении дозы доминантных генов обусловливает одинаковую выраженность признака (треугольная форма плода) независимо от их количества в генотипе.
К взаимодействию неаллельных генов относят также явление плейотропии — множественного действия гена, влияния его на развитие нескольких признаков. Плейотропное действие генов является результатом серьезного нарушения обмена веществ, обусловленного мутантной структурой данного гена.
Так, например, ирландские коровы породы декстер отличаются от близкой по происхождению породы керри укороченными ногами и головой, но одновременно лучшими мясными качествами и способностью к откорму. При скрещивании коров и быков породы декстер 25% телят имеют признаки породы керри, 50% сходны с породой декстер, а в остальных 25% случаев наблюдаются выкидыши уродливых бульдогообразных телят. Генетический анализ позволил установить, что причиной гибели части потомства является переход в гомозиготное состояние доминантной мутации, вызывающей недоразвитие гипофиза. В гетерозиготе этот ген приводит к появлению доминантных признаков коротконогости, короткоголовости и повышенной способности к отложению жира. В гомозиготе этот ген имеет летальный эффект, т.е. в отношении гибели потомства он ведет себя как рецессивный ген.
Летальный эффект при переходе в гомозиготное состояние характерен для многих плейотропных мутаций. Так, у лисиц доминантные гены, контролирующие платиновую и беломордую окраски меха, не оказывающие летального действия в гетерозиготе, вызывают гибель гомозиготных зародышей на ранней стадии развития. Аналогичная ситуация имеет место при наследовании серой окраски шерсти у овец породы ширази и недоразвития чешуи у зеркального карпа. Летальный эффект мутаций приводит к тому, что животные этих пород могут быть только гетерозиготными и при внутрипородных скрещиваниях дают расщепление в соотношении 2 мутанта: 1 норма.

F 1
F 1: 2 плат. : 1 черн.
Однако большинство летальных генов рецессивны, и гетерозиготные по ним особи имеют нормальный фенотип. О наличии у родителей таких генов можно судить по появлению в потомстве гомозиготных по ним уродов, абортусов и мертворожденных. Чаще всего подобное наблюдается в близкородственных скрещиваниях, где родители обладают сходными генотипами, и шансы перехода вредных мутаций в гомозиготное состояние достаточно высоки.
Плейотропные гены с летальным эффектом есть у дрозофилы. Так, доминантные гены Curly — загнутые вверх крылья, Star — звездчатые глаза, Notch — зазубренный край крыла и ряд других в гомозиготном состоянии вызывают гибель мух на ранних стадиях развития.
Известная рецессивная мутация white , впервые обнаруженная и изученная Т. Морганом, также имеет плейотропный эффект. В гомозиготном состоянии этот ген блокирует синтез глазных пигментов (белые глаза), снижает жизнеспособность и плодовитость мух и видоизменяет форму семенников у самцов.
У человека примером плейотропии служит болезнь Марфана (синдром паучьих пальцев, или арахнодактилия), которая вызывается доминантным геном, вызывающим усиленный рост пальцев. Одновременно он определяет аномалии хрусталика глаза и порок сердца. Болезнь протекает на фоне повышения интеллекта, в связи с чем ее называют болезнью великих людей. Ею страдали А. Линкольн, Н. Паганини.
Плейотропный эффект гена, по всей видимости, лежит в основе коррелятивной изменчивости, при которой изменение одного признака влечет за собой изменение других.
К взаимодействию неаллельных генов следует отнести также влияние генов-модификаторов, которые ослабляют или усиливают функцию основного структурного гена, контролирующего развитие признака. У дрозофилы известны гены-модификаторы, модифицирующие процесс жилкования крыльев. Известно не менее трех генов-модификаторов, влияющих на количество красного пигмента в волосе крупного рогатого скота, в результате чего окраска шерсти у разных пород колеблется от вишневой до палевой. У человека гены-модификаторы изменяют окраску глаз, усиливая или ослабляя ее интенсивность. Их действием объясняется разная окраска глаз у одного человека.
Существование явления взаимодействия генов привело к появлению таких понятий, как “генотипическая среда” и “генный баланс”. Под генотипической средой подразумевается то окружение, в которое попадает вновь возникающая мутация, т.е. весь комплекс генов, имеющихся в данном генотипе. Понятие “генный баланс” касается соотношения и взаимодействия между собой генов, влияющих на развитие признака. Обычно гены обозначают названием признака, возникающего при мутации. На самом же деле проявление этого признака часто является результатом нарушения функции гена под влиянием других генов (супрессоров, модификаторов и др.). Чем сложнее генетический контроль признака, чем больше генов участвуют в его развитии, тем выше наследственная изменчивость, так как мутация любого гена нарушает генный баланс и приводит к изменению признака. Следовательно, для нормального развития особи необходимо не только присутствие генов в генотипе, но и осуществление всего комплекса межаллельных и неаллельных взаимодействий.
Многочисленные опыты подтвердили правильность установленных Менделем закономерностей. Вместе с тем, появились факты, показывающие, что полученные Менделем числовые соотношения при расщеплении гибридного поколения соблюдались не всегда. Это указывало на то, что взаимоотношения между генами и признаками носят более сложный характер. Выяснилось: один и тот же ген может оказывать влияние на развитие нескольких признаков; один и тот же признак может развиваться под влиянием многих генов .
Следует отметить, что взаимодействие генов имеет биохимическую природу, то есть взаимодействуют друг с другом не гены, а их продукты. Продуктом эукариотического гена может быть или полипептид, или тРНК, или рРНК.
ВИДЫ ВЗАИМОДЕЙСТВИЯ АЛЛЕЛЬНЫХ ГЕНОВ
Различают полное доминирование, неполное доминирование, кодоминирование, аллельное исключение.
Аллельными генами называются гены, расположенные в идентичных локусах гомологичных хромосом. Ген может иметь одну, две и более молекулярных форм. Появление второй и последующих молекулярных форм является следствием мутации гена. Если ген имеет три и более молекулярных форм, говорят о множественном аллелизме . Из всего множества молекулярных форм у одного организма могут присутствовать только две, что объясняется парностью хромосом.
Полное доминирование
Полное доминирование — это вид взаимодействия аллельных генов, при котором фенотип гетерозигот не отличается от фенотипа гомозигот по доминанте, то есть в фенотипе гетерозигот присутствует продукт доминантного гена. Полное доминирование широко распространено в природе, имеет место при наследовании, например, окраски и формы семян гороха, цвета глаз и цвета волос у человека, резус-антигена и мн. др.
Наличие резус-антигена (резус-фактора) эритроцитов обусловливается доминантным геном Rh . То есть генотип резус-положительного человека может быть двух видов: или RhRh , или Rhrh ; генотип резус-отрицательного человека — rhrh . Если, например, мать — резус-отрицательная, а отец резус-положительный и гетерозиготен по этому признаку, то при данном типе брака с одинаковой вероятностью может родиться как резус-положительный, так и резус-отрицательный ребенок.
Между резус-положительным плодом и резус-отрицательной матерью может возникнуть резус-конфликт.

Так называется вид взаимодействия аллельных генов, при котором фенотип гетерозигот отличается как от фенотипа гомозигот по доминанте, так и от фенотипа гомозигот по рецессиву и имеет среднее (промежуточное) значение между ними. Имеет место при наследовании окраски околоцветника ночной красавицы, львиного зева, окраски шерсти морских свинок и пр.
Сам Мендель столкнулся с неполным доминированием, когда скрещивал крупнолистный сорт гороха с мелколистным. Гибриды первого поколения не повторяли признак ни одного из родительских растений, они имели листья средней величины.
При скрещивании гомозиготных красноплодных и белоплодных сортов земляники все первое поколение гибридов имеет розовые плоды. При скрещивании этих гибридов друг с другом получаем: по фенотипу — 1/4 красноплодных, 2/4 розовоплодных и 1/4 белоплодных растений, по генотипу — 1/4 АА , 1/2 Аа , 1/4 аа (и по фенотипу, и по генотипу соотношение 1:2:1). Соответствие расщепления по генотипу расщеплению по фенотипу является характерным при неполном доминировании, так как гетерозиготы фенотипически отличаются от гомозигот.
Кодоминирование
Кодоминирование — вид взаимодействия аллельных генов, при котором фенотип гетерозигот отличается как от фенотипа гомозигот по доминанте, так и от фенотипа гомозигот по рецессиву, и в фенотипе гетерозигот присутствуют продукты обоих генов. Имеет место при формировании, например, IV группы крови системы (АВ0) у человека.
Для того чтобы представить, как происходит наследование групп крови у человека, можно посмотреть, рождение детей с какой группой крови возможно у родителей, имеющих один — вторую, другой — третью группы крови и являющихся гетерозиготными по этому признаку.
| Р | ♀I A i 0
II (A) |
× | ♂I B i 0
III (B) |
||
| Типы гамет | I A | i 0 | I B | i 0 | |
| F | i 0 i 0
I (0) 25% |
I A i 0
II (A) 25% |
I B i 0
III (B) 25% |
I A I B
IV (AB) 25% |
|
Аллельное исключение
Аллельным исключением называется отсутствие или инактивация одного из пары генов; в этом случае в фенотипе присутствует продукт другого гена (гемизиготность, делеция, гетерохроматизация участка хромосомы, в котором находится нужный ген).
ВИДЫ ВЗАИМОДЕЙСТВИЯ НЕАЛЛЕЛЬНЫХ ГЕНОВ
Комплементарность, эпистаз, полимерия.
Неаллельные гены — гены, расположенные или в неидентичных локусах гомологичных хромосом, или в разных парах гомологичных хромосом.
Наука обрела обширную базу новых исследований субстрата эволюции - генетического кода. Именно в нем закладывается информация обо всех прошедших и предстоящих изменениях для развития организма.
Соотношение наследственности и изменчивости позволяет сохранять только лучшие качества, а взамен неудачных приобретать новые, совершенствуя строение и способствуя победе в естественном отборе.
Основные понятия генетики
В современной генетике за основу принята хромосомная теория наследования, согласно которой главным морфологическим субстратом является хромосома - структура из конденсированного комплекса ДНК (хроматина), с которого в процессе синтеза белков и считывается информация.

Генетика базируется на нескольких понятиях: ген (участок ДНК, кодирующий конкретный единичный признак), (совокупности генов и признаков организма), гаметы (половые клетки с одинарным набором хромосом) и зиготы (клетки с диплоидным набором).
Гены, в свою очередь, классифицируются на доминантные (А) и рецессивные (а) в зависимости от преобладания одного признака над другим, аллельные (А и а) и неаллельные гены (А и В). Аллельные располагаются на одинаковых участках хромосом и кодируют один признак. Неаллельные гены абсолютно противоположны им: располагаются на разных участках и кодируют разные признаки. Однако, несмотря на это, неаллельные гены имеют способность взаимодействовать друг с другом, порождая развитие совершенно новых признаков. По качественному составу аллельных генов организмы можно разделить на гомо- и гетерозиготные: в первом случае гены одинаковы (АА, аа), в другом - различны (Аа).
Механизм и схемы взаимодействия генов
Формы между собой изучал американский генетик Т.Х.Морган. Результаты своих исследований он изложил в Согласно ей гены, включенные в состав одной хромосомы, наследуются вместе. Такие гены получили название сцепленных и образуют т.н. группы сцепления. В свою очередь, внутри этих групп также происходит рекомбинация генов путем кроссинговера - обмена хромосом различными участками между собой. При этом абсолютно логичным и доказанным является тот факт, что гены, расположенные непосредственно друг за другом, разделению в процессе кроссинговера не подвергаются и наследуются совместно.

Если же между генами имеется расстояние, то вероятность разделения существует - данное явление получило название «неполное сцепление генов». Если говорить об этом более подробно, то взаимодействие аллельных генов между собой происходит по трем простым схемам: с получением чистого доминантного признака, неполное доминирование с получением промежуточного признака и кодоминирование с наследованием обоих признаков. Неаллельные гены же наследуются сложнее: по схемам комплементарности, полимерии или эпистаза. При этом наследоваться будут оба признака, но в разной мере.
Основные формы взаимодействия неалельных генов – комплементарность, эпистаз и полимерия. Они преимущественно видоизменяют классическую формулу расщепления за фенотипом, установленную Г. Менделем для дигибридного скрещивания (9:3:3:1).
Комплементарность (лат. complementum – дополнения). Комплементарными, или взаимодополняющими, называются неаллельные гены, которые поодиночке не проявляют своего действия, но при одновременном наличии в генотипе предопределяют развитие нового признака. У душистого горошка окраски цветков обусловлена двумя доминантными неаллельными генами, из них один ген (А) обеспечивает синтез бесцветного субстрата, другой (В) – синтез пигмента. Поэтому при скрещивании растений с белыми цветками (ААbb х ааВВ) все растения в первом поколении F1 (АаВb) имеют окрашенные цветки, а во втором поколении F2 происходит расщепления за фенотипом в соотношении 9:7, где 9/16 растений имеют окрашенные цветки и 7/16 – неокрашенные.
У человека нормальный слух обусловлен комплементарным взаимодействием двух доминантных неаллельных генов D и Е, из них один определяет развитие завитка, другой – слухового нерва. Люди с генотипами D–Е– имеют нормальный слух, с генотипами D–ее и ddЕ– – глухие. В браке, где родители глухие (DDee ´ ddEE), все дети будут иметь нормальный слух (DdEe).
Епистаз – взаимодействие неаллельных генов, при котором один ген подавляет действие другого, неаллельного, гена. Первый ген называется эпистатическим, или супрессором (ингибитором), другой, неаллельний, ген – гипостатическим. Если эпистатический ген – доминантный, эпистаз называют доминантным (А>В). И, наоборот, если эпистатический ген рецессивный, эпистаз – рецессивный (аа>В или аа >вв). Взаимодействие генов при эпистазе противоположно комплементарности.
Пример доминантного эпистаза . У кур доминантный аллель С одного гена обуславливает развитие окраски перья, но доминантный аллель І другого гена является его супрессором. Поэтому куры с генотипом І–С– – белые, а с генотипами ііСС и ііСс – окрашенные. В скрещивании белых кур (ІІСС х іісс) гибриды первого поколения F1 окажутся белыми, но при скрещивании F1 между собой во втором поколении F2 состоится расщепления за фенотипом в соотношении 13:3. Из 16 особей 3 будут окрашены (ііСС и ііСс), так как в них отсутствует доминантный ген-супрессор и есть доминантный ген окраски. Другие 13 особей будут белыми.
Примером рецесивного эпистаза может быть бомбейский феномен – необыкновенное наследование групп крови системы АВО, впервые выявленное в одной индийской семье. В семье, где отец имел группу крови І (О), а иметь – ІІІ (В), родилась девочка с группой І (О), она вступила в брак с мужчиной с группой крови ІІ(А) и у них родилось две девочки: одна из группой крови ІV (АВ), другая – с І (О). Рождение девочки с ІV (АВ) группой крови в семье, где отец имел ІІ (А), а мама – І (О) было необыкновенным. Генетики объяснили этот феномен так: девочка с группой ІV (АВ) унаследовала аллель ІА от отца, а аллель ІВ – от матери, но у матери аллель ІВ фенотипически не проявлялся, так как в ее генотипе присутствовал редкий рецессивний эпистатический ген s в гомозиготном состоянии, который спровоцировал фенотипичное проявление аллеля ІВ.
Гипостаз – взаимодействие неалельных генов, при котором доминантный ген одной аллельной пары подавляется эпистатическим геном из другой аллельной пары. Если ген А подавляет ген В (А>В), то по отношению к гену В взаимодействие неаллельных генов называется гипостазом, а по отношению к гену А – эпистазом.
Полимерия – взаимодействие неаллельных генов, при котором один и и тот же признак контролируют несколько доминантных неаллельных генов, которые действуют на этот признак однозначно, в равной степени, усиливая его проявление. Такие однозначные гены называют полимерными (множественными, полигенами) и их обозначают одной буквой латинского алфавита, но с разными цифровыми индексами. Например, доминантные полимерные гены – А1, А2, А3 и т.д., рецессивные – а1, а2, а3 и т.д. Соответственно обозначают генотипы – А1А1А2А2А3А3, а1а1а2а2а3а3. Признаки, которые контролируются полигенами, называют полигенными, а наследования этих признаков – полигенным, в отличие от моногенного, где признак контролируется одним геном. Явление полимерии впервые описал в 1908 г. шведский генетик Г. Нильсон-Эле при изучении наследования цвета зерна пшеницы.
Полимерия бывает кумулятивной и некумулятивной . При кумулятивной полимерии каждый ген в отдельности имеет слабое действие (слабую дозу), но количество доз всех генов в конечном результате суммируется, так что степень выражения признака зависит от числа доминантных аллелей. За типом полимерии у человека наследуются рост, масса тела, цвет кожи, умственные способности, величина артериального давления. Так, пигментация кожи у человека определяется 4-6 парами полимерных генов. В генотипе коренных жителей Африки имеются преимущественно доминантные аллели (Р1Р1Р2Р2Р3Р3Р4Р4), у представителей европеоидной рассы – рецессивные (p1p1p2p2p3p3p4p4). От брака темнокожего и белой женщины рождаются дети с промежуточным цветом кожи – мулаты (Р1р1P2р2P3р3P4р4). Если супруги – мулаты, то возможное рождение детей с пигментацией кожи от максимально светлой к максимально темной.
Полигенно в типичных случаях наследуются количественные признаки. Тем не менее в природе существуют примеры полигенного наследования качественных признаков, когда конечный результат не зависит от числа доминантных аллелей в генотипе – признак или проявляется, или не проявляется (некумулятивная полимерия).
Плейотропия – способность одного гена контролировать несколько признаков (множественное действие гена). Так, синдром Марфана в типичных случаях характеризуется триадой признаков: подвывихом хрусталика глаза, пороками сердца, удлинением костей пальцев рук и ног (арахнодактилия – паучьи пальцы). Этот комплекс признаков контролируется одним аутосомно-доминантным геном, который вызывает нарушения развития соединительной ткани.